


La nutrición deportiva es la rama de la nutrición especializada en las personas que practican deporte, mediante la cual se aconseja, se guía y se educa al deportista a saber qué, cuándo y por qué se deben comer y beber ciertos alimentos u otros según la ocasión.
Para ponernos en contexto vamos a repasar algunos conceptos básicos y aplicarlos al AYUNO INTERMITENTE.
Es fundamental tener conocimientos sobre el metabolismo y la bioenergética.
El movimiento humano, tanto en la vida diaria como en el ámbito deportivo, se caracteriza por ser un evento energético.
La ciencia que estudia los principios que limitan el intercambio de energía se conoce como termodinámica o energética.

Para describir la energía en el cuerpo humano se deben tener presente dos cosas.
- En primer lugar, la energía no se crea sino que se obtiene en una forma y se convierte en otra.
- En segundo lugar, los procesos de conversión de energía son relativamente ineficientes, y la mayoría de la energía se libera en una forma no utilizable: calor (Brooks et al., 2013).
1. El Metabolismo

El metabolismo abarca todas las reacciones químicas que se llevan a cabo en el cuerpo; incluye la degradación de moléculas complejas en estructuras más pequeñas, es decir, catabolismo y la síntesis de moléculas más complejas a partir de otras más simples y de menor tamaño, es decir, anabolismo. (Tortora & Derrickson, 2008). En forma más simple, puede ser definido como la suma de todos los procesos que ocurren en un organismo vivo (Brooks et al., 2013).
Las reacciones químicas suceden en el organismo cuando se forman o se destruyen enlaces químicos entre las sustancias. Las enzimas funcionan como catalizadores para aumentar la velocidad en las reacciones químicas.
El metabolismo podría considerarse como un proceso que mantiene el equilibrio energético entre las reacciones anabólicas y catabólicas (Tortora& Derrickson, 2008).
Durante las reacciones anabólicas se consume más energía de la que se produce durante su desarrollo; esta energía proviene de las reacciones catabólicas. A su vez, las reacciones catabólicas liberan la energía almacenada en las moléculas que degradan; esta energía se transfiere al adenosintrifosfato (ATP), y luego se usa para abastecer de energía a las reacciones anabólicas.
El calor producido por todas estas reacciones químicas es lo que se denomina tasa metabólica, y está reflejada en la tasa de producción de calor. Todas las reacciones dependen, en última instancia, de las reacciones de oxidación biológica; por lo tanto, la evaluación del consumo de oxígeno es una buena aproximación de la tasa de producción de calor o tasa metabólica (Brooks et al., 2013).

Aquí te dejo un video del Profesor Matías Rodríguez que es muy didáctico y que te va a servir para comprender a la perfección de qué se trata el Metabolismo.
Disfrutalo!!

Antes de continuar profundizando diferentes aspectos de la bioenergética, es preciso definir algunos términos importantes (Brooks et al., 2013):
· Energía: es la capacidad de realizar un trabajo.
· Trabajo: es el producto de una fuerza dada, actuando durante una distancia. Esta es una definición mecánica. En las células es más común que se realice un trabajo biológico (químico y eléctrico), que mecánico. Sin embargo, es posible intercambiar y convertir energía de una forma a otra.
· Potencia: tasa de producción de un trabajo.
· Sistema: unidad funcionalmente organizada. Los sistemas pueden variar desde el nivel microscópico (mitocondria) hasta el nivel corporal total.
· Turnover: en el organismo, las moléculas de energía son constantemente utilizadas y restauradas. El turnover se refiere a la tasa de renovación, en la cual un estado estable del metabolismo refleja que la tasa de utilización es equivalente a la de restauración.
2. Producción de energía mecánica y sistemas energéticos

El estudio de la energía y los principios que regulan su intercambio comenzó en el Siglo XIX y se denominó Termodinámica.
Hoy en día, el término más adecuado y más frecuentemente utilizado es Energética.
Existen seis formas básicas de energía: térmica, química, mecánica, eléctrica, lumínica y atómica.
Las máquinas mecánicas convierten energía en calor, el cual es convertido en energía mecánica.
En cambio, las máquinas biológicas (el cuerpo humano) no tienen la capacidad de convertir el calor en otro tipo de energía. En los sistemas biológicos, el calor es un componente esencial, pero inútil, de las reacciones en las cuales otras formas de trabajo se han realizado (Brooks et al., 2013).
Los mecanismos para la conversión de energía están presentes en todas las células. Ellas necesitan de una sustancia que sea capaz de recibir la energía que se libera en numerosas reacciones y, al mismo tiempo, también que sea capaz de proporcionar energía en las reacciones que precisan de ella.
En nuestras células esa sustancia es casi siempre el ATP, por ello, frecuentemente se lo reconoce como un intermediario común de energía (Brooks et al., 2013).
El ATP pertenece al grupo de los nucleótidos.
Su estructura está conformada por una base nitrogenada (adenina), un azúcar (ribosa) y 3 fosfatos, siendo el último de estos el que habitualmente se denomina de “alta energía”.
La reacción por la cual el ATP es dividido en adenosindifosfato (ADP) para liberar energía involucra agua, por lo que este tipo de reacciones son llamadas hidrólisis (que significa: dividido o escindido por el agua).

Por lo tanto, el ATP puede ser considerado como un intermediario químico común usado para brindar energía al trabajo muscular y otras formas de trabajo celular.
Sin embargo, la concentración de ATP en las células musculares es baja.
Se estima que esta cantidad puede abastecer a un trabajo muscular de máxima intensidad solo durante 2 segundos (Platonov, 2001).
De todos modos, las células están preparadas para mantener la concentración celular de ATP relativamente constante, en varias tasas de uso (tasas de turnover); esto es lo que se conoce como homeostasis del ATP (Brooks et al., 2013).
El músculo cuenta con tres fuentes de energía (o sistemas de producción de energía) que permiten dar soporte a la homeostasis del ATP: inmediatas, no oxidativas, oxidativas. (Brooks et al., 2013).

3. Fuentes inmediatas de energía

Las fuentes inmediatas de energía en el músculo están conformada por tres componentes.
- En primer lugar, la reserva de ATP. La reacción que degrada el ATP es catalizada por la enzima ATPasa, y se puede resumir de la siguiente manera:
ATP + H2O = ADP + Pi + Energía
- La segunda fuente de energía celular inmediata es la Fosfocreatina (PCr), que proporciona una reserva energética de fosfato para regenerar el ATP consumido durante la contracción muscular. La interacción entre el ADP, la PCr y el ATP degradado es catalizada por la enzima Creatinquinasa, y se puede resumir en:
ADP + PCr = ATP + Cr
- La tercer fuente inmediata de energía celular involucra a la enzima Adenilato Quinasa, que en el músculo se suele llamar Mioquinasa, y cataliza esta reacción:
ADP + ADP = ATP + AMP
El AMP es importante, ya que su presencia en la célula muscular es una señal importante para la activación de los mecanismos de restauración de ADP en ATP (Brooks et al., 2013).
El ATP y la PCr, que conjuntamente suelen ser llamados fosfágenos, constituyen una reserva de energía muy importante, por estar disponibles en forma inmediata, pero que no puede sostener su trabajo más allá de unos pocos segundos (5-15 s), necesitando de la ayuda de otras fuentes de energía (Brooks et al., 2013).
- Fuentes de energía no oxidativas (glucolíticas)
Los combustibles utilizados por este sistema energético son la glucosa y el glucógeno, y los procesos por los cuales se obtiene energía a partir de ellos se denominan glucólisis y glucogenólisis, respectivamente.
La glucólisis podría ser resumida en la siguiente ecuación (Brooks et al., 2013):
Glucosa = 2 ATP + 2 lactato- + 2 H+
Cuantitativamente, la energía disponible por las fuentes no oxidativas es muy superior a la obtenida por las fuentes inmediatas.
Sin embargo, aunque se combine la energía obtenida por estas dos fuentes, la cantidad obtenida es una fracción muy pequeña comparada con la energía disponible a través de las fuentes oxidativas.
Por lo tanto, actividades musculares intensas con una duración mayor a 30 s no pueden ser mantenidas sin la ayuda del metabolismo oxidativo.
Además, el metabolismo oxidativo es necesario para la restauración de los sistemas inmediatos y no oxidativos luego del ejercicio intenso (Brooks et al., 2013).
- Fuentes de energía oxidativas
Los potenciales combustibles del sistema energético oxidativo incluyen los CHO, las grasas y algunos aminoácidos (AA) (Figura 4). Si bien la glucosa puede ser utilizada a través de mecanismos glucolíticos (ver ecuación anterior), las reacciones oxidativas permiten extraer más energía de una molécula de glucosa:
Glucosa + O2 = 36 ATP + CO2 + H2O
Esto se debe a que el metabolismo oxidativo permite un catabolismo más completo de la molécula de glucosa.
Los ácidos grasos (AG), al ser catabolizados por el metabolismo oxidativo, liberan una cantidad de energía mucho mayor que la glucosa.
Por ejemplo, si se oxida una molécula de palmitato (un AG de tamaño medio y bastante común) se obtiene:
Palmitato + O2 = 129 ATP + CO2 + H2O
Los aminoácidos (AA) también pueden seguir la vía oxidativa pero para ello, previamente, el residuo que contiene el nitrógeno (N2) debe ser removido.
Esto generalmente es realizado a través del intercambio del N2 con otro compuesto (transaminación), o bien a través de un proceso único de remoción del N2 (desaminación oxidativa). De todos modos, los AA no suelen ser utilizados para la producción de energía en el músculo, y solo dan cuenta de una pequeña fracción del ATP sintetizado (Brooks et al., 2013).

4. Cuantificación del gasto energético de la actividad física

Una cuestión que muchas veces se pasa por alto es que el término actividad física (AF) no es sinónimo de gasto energético (GE). La AF es una conducta caracterizada por el movimiento corporal, consecuencia de la acción muscular, y que resulta en un GE.
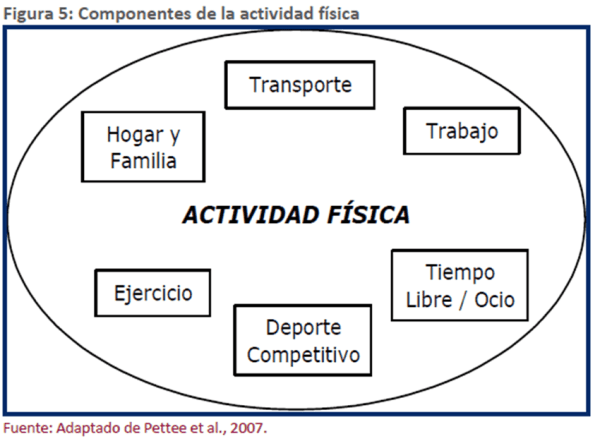
Se pueden reconocer varios tipos o categorías de AF (Figura 5) que, en ocasiones, incluso se superponen, dependiendo del objetivo con el cual se realice la categorización.
Por ejemplo, una caminata rápida puede ser un medio de transporte, o bien formar parte de un programa de ejercicio planificado. Debido a esta superposición de las subcategorías de la AF, es muy complicada su medición como categorías independientes (Pettee, Tudor-Locke, & Ainsworth, 2007).
Otra categorización de la AF podría basarse en la intensidad con la que se realiza, es decir la tasa del GE atribuida a la actividad específica (Ainsworth et al., 1993; Ainsworth et al., 2000; Pate et al., 1995).
La AF generalmente se tipifica en base a su frecuencia (p. ej., cantidad de veces por semana) y la duración (p. ej., minutos de duración de cada sesión).
Pero el GE refleja el costo metabólico de una AF específica, y es el producto de la frecuencia, la duración y la intensidad de dicha actividad.
Otro punto importante referido a la estimación del GE relacionado a una actividad es la posibilidad de usar escalas donde la expresión de la intensidad o el costo energético puede hacerse de manera absoluta (ejemplo, kcal/kg/hora) o relativa (como % de la máxima capacidad actual).
A pesar de que varios factores pueden afectar el GE en una escala relativa (por ej., la edad, el peso corporal, el nivel de rendimiento), si asumimos una eficiencia mecánica relativamente constante en el ser humano (~23%) para desarrollar trabajo mecánico, entonces el GE absoluto es generalmente constante para una actividad determinada.
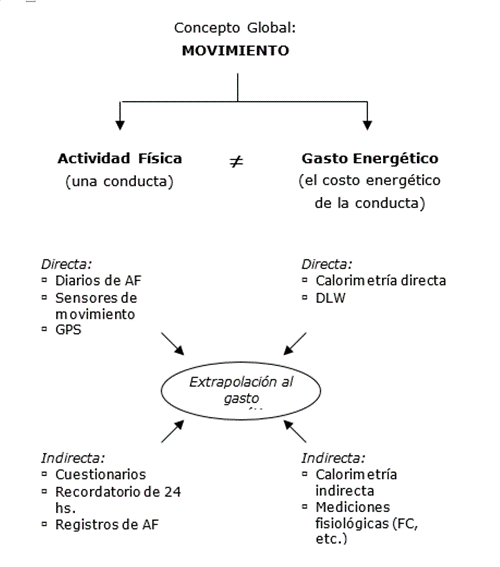
Para facilitar la comprensión de la evaluación de la AF y el GE podría ser útil tener un marco conceptual en el que el concepto central de interés sea movimiento.
A su vez, este concepto se puede operacionalizar en dos variables mensurables: AF y GE . Ambas variables se pueden evaluar a través de mediciones directas e indirectas.
Sin embargo, generalmente se trata de extrapolar los resultados de estas mediciones a alguna unidad de medida del GE (kcal o kJ) debido a su relación con la ingesta de energía y, en última instancia, su efecto sobre el BE.
En resumen, para poder equilibrar adecuadamente la ingesta energética de un atleta con su gasto energético son necesarias medidas válidas que permitan cuantificar sus patrones de AF y el GE asociado.
La importancia de lograr cubrir adecuadamente las necesidades energéticas de los atletas se basa en que la ingesta de energía adecuada es esencial para el mantenimiento de la masa magra, la función inmune y reproductiva, así como para una óptima performance deportiva (American College of Sports Medicine, 2000).
Además, un déficit crónico en la ingesta de energía ha sido considerado como uno de los factores causales potenciales del sobreentrenamiento (Leutholtz & Kreider, 2001).
5. Reservas de energía

Los sustratos de los que se vale nuestro cuerpo para producir ATP, siendo la vía oxidativa la principal (salvo cuando se trata de esfuerzos intensos y breves), son los carbohidratos (CHO), las grasas y las proteínas.
De todos modos, si bien las proteínas pueden ser utilizadas como combustible, sus principales funciones son la estructural y la reguladora, por lo que no se consideran sustratos energéticos importantes. Esto convierte a los CHO y las grasas en los principales sustratos energéticos para el músculo esquelético durante el ejercicio (Figura 7) (Coyle, 1997).
Los triglicéridos (TG) almacenados dentro de los adipocitos del tejido adiposo corporal pueden representar unos 5 a 10 kg en hombres y mujeres que posean entre un 10% a 30% de grasa corporal.
Estos TG de reserva son hidrolizados, a través de la lipólisis, a glicerol y ácidos grasos libres (AGL). Los AGL por tratarse de moléculas hidrofóbicas (por ejemplo, sustancias que no tienen afinidad por el agua) deben unirse a la albúmina para poder ser transportados en la sangre hasta, por ejemplo, el músculo esquelético (Figura 7).
 5.
5.

Responses
Lo siento, debes estar conectado para publicar un comentario.
Responses